Vestibular System Overview
•Download as PPT, PDF•
0 likes•4 views

The vestibular system detects motion and orientation of the head to control posture, balance, and eye movement. It contains the maculae in the saccule and utricle, which detect linear acceleration, and semicircular canals, which detect angular acceleration. The vestibular system provides inputs that allow for anticipatory postural adjustments and learned adaptive control of balance and posture.
![Overview of Vestibular System ,[object Object],[object Object]](data:image/gif;base64,R0lGODlhAQABAIAAAAAAAP///yH5BAEAAAAALAAAAAABAAEAAAIBRAA7)
Recommended
More Related Content
What's hot
What's hot (20)
Similar to Vestibular System Overview
Similar to Vestibular System Overview (20)
More from PS Deb
More from PS Deb (20)
Recently uploaded
Recently uploaded (20)
Vestibular System Overview
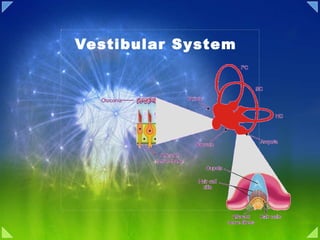
- 5. Macule and Hair Cell
- 6. Vestibular transduction of Mechanical Stimuli
- 7. Morphological polarization of hair cells in the utricular and saccular maculae
- 8. Forces acting on the head and the resulting displacement of the otolithic membrane of the utricular macula
- 9. Response of a vestibular nerve axon from an otolith organ
- 12. The Semicircular Canals Detect Angular Accelerations
- 14. The ampulla is responsible for the change in rotational movement
- 21. Postural adjustments require anticipatory motor actions.
- 22. Anticipatory maintenance of body posture
- 23. Anticipatory motor action in response to postural disturbance
- 24. Appropriate anticipatory responses to postural disturbances can be learned
- 25. Adaptive Postural Control Is Learned During Locomotion
- 27. The vestibular contribution to postural adjustment depends on whether the primary perturbation is to the feet or the head
Editor's Notes
- Michael E. Goldberg A. J. Hudspeth AIRPLANES AND SUBMARINES navigate in three dimensions using sophisticated guidance systems that register every acceleration and turn. Laser gyroscopes and computers have afforded such navigational aids unprecedented precision. Yet the principles of inertial guidance are ancient—vertebrates have used analogous systems for 500 million years, and invertebrates for still longer. The vestibular system is designed to answer two of the questions basic to human life: “Which way is up?” and “Where am I going?” It does so by measuring linear and angular acceleration of the head through an ensemble of five sensory organs in the inner ear (the membranous or vestibular labyrinth). Acceleration of the head deflects hair bundles attached to hair cells in the vestibular labyrinth. This distortion changes membrane potential and transmitter release patterns in these cells, thereby affecting the discharge patterns of the vestibular neurons that innervate them. In turn, the vestibular neurons carry head velocity and acceleration signals to the vestibular nuclei in the brain stem. This information helps us maintain balance and influences how we perceive space. In this chapter we consider how the hair cells of the vestibular apparatus generate the primary signals for head acceleration and how the brain integrates these signals. The early origin of the vestibular system is seen in the outputs of the labyrinthine receptors, which flow to vestibular nuclei that occupy a prominent position in the brain stem. As we shall learn in the next chapter, various components of the vestibular system subserve a variety of postural reflexes, including those that make possible upright, bipedal posture. Finally, through pathways involving the ocular motor nuclei and cerebellum, the vestibular system also controls reflex eye movements that stabilize retinal images despite head and body motions.
- It is the primary organ of equilibrium and thus plays a major role in the subjective sensation of motion and spatial orientation. Vestibular input to areas of the nervous system involved in motor control elicits adjustments of muscle activity and body position to allow for upright posture . 3. Vestibular input to regions of the nervous system controlling eye movements helps stabilize the eyes in space during head movements. This reduces the movement of the image of a fixed object on the retina. The vestibular system detects motion of the head-in-space and in turn generates reflexes that are crucial for our daily activities, such as stabilizing the visual axis (gaze) and maintaining head and body posture. In addition, the vestibular system provides us with our subjective sense of movement and orientation in space. The vestibular sensory organs are located in the petrous part of the temporal bone in close proximity to the cochlea , the auditory sensory organ. Although the vestibular system was only recognized as a separate entity from the auditory portion of the inner ear in the middle of the 19th century, it is phylogenetically the oldest part of the inner ear. The vestibular system is comprised of two types of sensors: the two otolith organs (the saccule and utricle), which sense linear acceleration (i.e., gravity and translational movements), and the three semicircular canals, which sense angular acceleration in three planes. The receptor cells of the otoliths and semicircular canals send signals through the vestibular nerve fibers to the neural structures that control eye movements , posture, and balance (Fig. 1 ). In contrast to the senses of vision and audition which can easily be understood by simply shutting our eyes or plugging our ears, the significance of vestibular function in our daily lives is more difficult to appreciate. When the system is functioning normally, we are usually unaware of a distinct sensation arising from vestibular activity since it is integrated with visual, proprioceptive and other sensory information such that combined experience leads to a sense of motion. In addition, the important contributions that the vestibular system normally makes to gaze stabilization and postural control are difficult to fully comprehend. For this reason, clinical studies have provided many important insights into function of the vestibular system. Perhaps the best known clinical case is a report by a physician JC in 1952, who suffered from complete vestibular loss following treatment with streptomycin (an ototoxic antibiotic) for leg infection. He described his symptoms following treatment, and commented that “By bracing my head between two of the metal bars at the head of the bed I found I could minimize the effect of the pulse beat that made the letters of the page jump and blur” and that [When walking] “in these corridors I had the peculiar sensation that I was inside a flexible tube, fixed at the end nearest me but swaying free at the far end.”
- located in the petrous portion of the temporal bone, called the bony labyrinth. Within this system are membranous tubes and chambers called the membranous labyrinth. The membranous labyrinth is the functional part of the vestibular apparatus. The top of Figure 55–9 shows the membranous labyrinth. It is composed mainly of the cochlea (ductus cochlearis); three semicircular canals; and two large chambers, the utricle and saccule. The cochlea is the major sensory organ for hearing (see Chapter 52) and has little to do with equilibrium. However, the semicircular canals, the utricle, and the saccule are all integral parts of the equilibrium mechanism. “ Maculae”—Sensory Organs of the Utricle and Saccule for Detecting Orientation of the Head with Respect to Gravity. Located on the inside surface of each utricle and saccule, shown in the top diagram of Figure 55–9, is a small sensory area slightly over 2 millimeters in diameter called a macula. The macula of the utricle lies mainly in the horizontal plane on the inferior surface of the utricle and plays an important role in determining orientation of the head when the head is upright. Conversely, the macula of the saccule is located mainly in a vertical plane and signals head orientation when the person is lying down. Each macula is covered by a gelatinous layer in which many small calcium carbonate crystals called statoconia are embedded .Also in the macula are thousands of hair cells, one of which is shown in Figure 55–10; these project cilia up into the gelatinous layer. The bases and sides of the hair cells synapse with sensory endings of the vestibular nerve. The calcified statoconia have a specific gravity two to three times the specific gravity of the surrounding fluid and tissues. The weight of the statoconia bends the cilia in the direction of gravitational pull. The Utricle and the Saccule Detect Linear Accelerations The simplest labyrinthine organs are the utricle (or utriculus ) and the saccule (or sacculus ), each of which consists of an ovoidal sac of membranous labyrinth about 3 mm in the longest dimension. The complement of hair cells in each organ is localized to a roughly elliptical patch, called the macula. The human utricle contains about 30,000 hair cells, while the saccule contains some 16,000. The hair bundle at the apex of each hair cell extends into the endolymphatic space of the utricle or saccule, where the bundle's top is attached to a gelatinous sheet, the otolithic membrane , that covers the entire sensory macula (Figure 40-3). Embedded within and lying on the otolithic membrane are fine, dense particles, the otoconia (“ear dust”); consisting of calcium carbonate in the form of the mineral calcite. Otoconia are typically 0.5-10 μm long and millions of these particles fill the endolymphatic cavities of the utricle and the saccule. Because of the prominence of otoconia, the utricle and saccule are named the otolithic organs. When the head undergoes linear acceleration the membranous labyrinth moves along as well because it is fixed to the skull. The otoconial mass, however, is free to shift within the receptor organ. Because of its inertia this mass lags behind movement of the head. The motion of the otoconia is communicated to the gelatinous otolithic membrane, which thus shifts with respect to the underlying epithelium. This motion in turn deflects the hair bundles that link the otolithic membrane to the macula, thus exciting an electrical response in the hair cells.
- Figure 14.2. The morphological polarization of vestibular hair cells and the polarization maps of the vestibular organs. (A) A cross section of hair cells shows that the kinocilia of a group of hair cells are all located on the same side of the hair cell. The arrow indicates the direction of deflection that depolarizes the hair cell. (B) View looking down on the hair bundles. (C) In the ampulla located at the base of each semicircular canal, the hair bundles are oriented in the same direction. In the sacculus and utricle, the striola divides the hair cells into populations with opposing hair bundle polarities.
- Each macula is covered by a gelatinous layer in which many small calcium carbonate crystals called statoconia are embedded .Also in the macula are thousands of hair cells, one of which is shown in Figure 55–10; these project cilia up into the gelatinous layer. The bases and sides of the hair cells synapse with sensory endings of the vestibular nerve. The calcified statoconia have a specific gravity two to three times the specific gravity of the surrounding fluid and tissues. The weight of the statoconia bends the cilia in the direction of gravitational pull. The Otolith Organs: The Utricle and Sacculus Displacements and linear accelerations of the head, such as those induced by tilting or translational movements (see Box A), are detected by the two otolith organs: the sacculus and the utricle. Both of these organs contain a sensory epithelium, the macula, which consists of hair cells and associated supporting cells. Overlying the hair cells and their hair bundles is a gelatinous layer, and above this is a fibrous structure, the otolithic membrane, in which are embedded crystals of calcium carbonate called otoconia (Figures 14.3 and 14.4A). The crystals give the otolith organs their name ( otolith is Greek for “ear stones”). The otoconia make the otolithic membrane considerably heavier than the structures and fluids surrounding it; thus, when the head tilts, gravity causes the membrane to shift relative to the sensory epithelium (Figure 14.4B). The resulting shearing motion between the otolithic membrane and the macula displaces the hair bundles, which are embedded in the lower, gelatinous surface of the membrane. This displacement of the hair bundles generates a receptor potential in the hair cells. A shearing motion between the macula and the otolithic membrane also occurs when the head undergoes linear accelerations (see Figure 14.5); the greater relative mass of the otolithic membrane causes it to lag behind the macula temporarily, leading to transient displacement of the hair bundle. The similar effects exerted on otolithic hair cells by certain head tilts and linear accelerations explains the perceptual equivalency of these different stimuli when visual feedback is absent, as occurs in the dark or when the eyes are closed. As already mentioned, the orientation of the hair cell bundles is organized relative to the striola, which demarcates the overlying layer of otoconia (see Figure 14.4A). The striola forms an axis of mirror symmetry such that hair cells on opposite sides of the striola have opposing morphological polarizations. Thus, a tilt along the axis of the striola will excite the hair cells on one side while inhibiting the hair cells on the other side. The saccular macula is oriented vertically and the utricular macula horizontally, with a continuous variation in the morphological polarization of the hair cells located in each macula (as shown in Figure 14.4C, where the arrows indicate the direction of movement that produces excitation). Inspection of the excitatory orientations in the maculae indicates that the utricle responds to movements of the head in the horizontal plane, such as sideways head tilts and rapid lateral displacements, whereas the sacculus responds to movements in the vertical plane (up-down and forward-backward movements in the sagittal plane). Note that the saccular and utricular maculae on one side of the head are mirror images of those on the other side. Thus, a tilt of the head to one side has opposite effects on corresponding hair cells of the two utricular maculae. This concept is important in understanding how the central connections of the vestibular periphery mediate the interaction of inputs from the two sides of the head (see next section). Directional Sensitivity of the Hair Cells—Kinocilium. Each hair cell has 50 to 70 small cilia called stereocilia, plus one large cilium, the kinocilium, as shown in Figure 55–10. The kinocilium is always located to one side, and the stereocilia become progressively shorter toward the other side of the cell. Minute filamentous attachments, almost invisible even to the electron microscope, connect the tip of each stereocilium to the next longer stereocilium and, finally, to the kinocilium. Because of these attachments, when the stereocilia and kinocilium bend in the direction of the kinocilium, the filamentous attachments tug in sequence on the stereocilia, pulling them outward from the cell body. This opens several hundred fluid channels in the neuronal cell membrane around the bases of the stereocilia, and these channels are capable of conducting large numbers of positive ions.Therefore, positive ions pour into the cell from the surrounding endolymphatic fluid, causing receptor membrane depolarization. Conversely, bending the pile of stereocilia in the opposite direction (backward to the kinocilium) reduces the tension on the attachments; this closes the ion channels, thus causing receptor hyperpolarization. Under normal resting conditions, the nerve fibers leading from the hair cells transmit continuous nerve impulses at a rate of about 100 per second. When the stereocilia are bent toward the kinocilium, the impulse traffic increases, often to several hundred per second; conversely, bending the cilia away from the kinocilium decreases the impulse traffic, often turning it off completely. Therefore, as the orientation of the head in space changes and the weight of the statoconia bends the cilia, appropriate signals are transmitted to the brain to control equilibrium. In each macula, each of the hair cells is oriented in a different direction so that some of the hair cells are stimulated when the head bends forward, some are stimulated when it bends backward, others are stimulated when it bends to one side, and so forth. Therefore, a different pattern of excitation occurs in the macular nerve fibers for each orientation of the head in the gravitational field. It is this “pattern” that apprises the brain of the head’s orientation in space. Figure 40-3 The utricle is organized to detect tilt of the head. The hair cells in the epithelium of the utricle have apical hair bundles that project into the otolithic membrane, a gelatinous material embedded with calcium carbonate stones (otoconia). The hair bundles are polarized, but not all cells are oriented in the same direction. The response of an individual hair cell in the utricle to a tilt of the head depends on the direction in which its hairs are bent by the gravitational force of the otoliths. When the head is tilted in the direction of the axis of polarity for a particular hair cell, that cell depolarizes and excites the afferent fiber. When the head is tilted in the opposite direction, the hair cell hyperpolarizes and inhibits the afferent fiber (see Figure 40-2). (Adapted from Iurato 1967.) Vestibular Hair Cells The vestibular hair cells, which, like cochlear hair cells, transduce minute displacements into behaviorally relevant receptor potentials, provide the basis for vestibular function. Vestibular and auditory hair cells are quite similar; a detailed description of hair cell structure and function has already been given in Chapter 13. As in the case of auditory hair cells, movement of the stereocilia toward the kinocilium in the vestibular end organs opens mechanically gated transduction channels located at the tips of the stereocilia, depolarizing the hair cell and causing neurotransmitter release onto (and excitation of) the vestibular nerve fibers. Movement of the stereocilia in the direction away from the kinocilium closes the channels, hyperpolarizing the hair cell and thus reducing vestibular nerve activity. The biphasic nature of the receptor potential means that some transduction channels are open in the absence of stimulation, with the result that hair cells tonically release transmitter, thereby generating considerable spontaneous activity in vestibular nerve fibers (Box B). One consequence of these spontaneous action potentials is that the firing rates of vestibular fibers can increase or decrease in a manner that faithfully mimics the receptor potentials produced by the hair cells. Importantly, the hair cell bundles in each vestibular organ have specific orientations (Figure 14.2). As a result, the organ as a whole is responsive to displacements in all directions. In a given semicircular canal, the hair cells in the ampulla are all polarized in the same direction (Figure 14.2C). In the utricle and sacculus, a specialized area called the striola divides the hair cells into two populations with opposing polarities (Figures 14.2C; see also Figure 14.4C). The directional polarization of the receptor surfaces is a basic principle of organization in the vestibular system, as will become apparent in the following descriptions of the individual vestibular organs Function of the Utricle and Saccule in the Maintenance of Static Equilibrium It is especially important that the hair cells are all oriented in different directions in the maculae of the utricles and saccules, so that with different positions of the head, different hair cells become stimulated.The “patterns” of stimulation of the different hair cells apprise the brain of the position of the head with respect to the pull of gravity. In turn, the vestibular, cerebellar, and reticular motor nerve systems of the brain excite appropriate postural muscles to maintain proper equilibrium.
- Figure 40-2 Hair cells in the vestibular labyrinth transduce mechanical stimuli into neural signals. At the apex of each cell is a hair bundle in which a number of stereocilia taper in length toward a single kinocilium. The membrane potential of the receptor cell depends on the direction in which the hair bundle is bent. Deflection toward the kinocilium causes the cell to depolarize and thus increases the rate of firing in the afferent fiber. Bending away from the kinocilium causes the cell to hyperpolarize, thus decreasing the afferent firing rate. (Adapted from Flock 1965.) During its development the labyrinth progresses from a simple sac to a complex of interconnected organs. However, its fundamental topological organization persists: Each organ originates as an epithelium-lined pouch that buds from the otic cyst, and the endolymphatic spaces within the several organs remain continuous in the adult. The endolymphatic spaces of the vestibular labyrinth are also connected to the scala media of the cochlea through the ductus reuniens. Among the epithelial cells lining the membranous labyrinth are five clusters of hair cells, one cluster in each receptor organ. Like the hair cells that mediate hearing in the cochlea, the hair cells of the vestibular labyrinth are endowed with hair bundles that transduce mechanical stimuli into receptor potentials. The general principles of mechanoelectrical transduction by hair cells (Chapter 31) also apply to these cells. Deflection of a hair bundle toward the kinocilium elicits a depolarization, which in turn enhances the release of synaptic transmitter. Deflection away from the kinocilium hyperpolarizes the hair cell and reduces neurotransmitter release (Figure 40-2).
- Figure 14.4. Morphological polarization of hair cells in the utricular and saccular maculae. (A) Cross section of the utricular macula showing hair bundles projecting into the gelatinous layer when the head is level. (B) Cross section of the utricular macula when the head is tilted. (C) Orientation of the utricular and saccular maculae in the head; arrows show orientation of the kinocilia, as in Figure 14.2 . The saccules on either side are oriented more or less vertically, and the utricles more or less horizontally. The striola is a structural landmark consisting of small otoconia arranged in a narrow trench that divides each otolith organ. In the utricular macula, the kinocilia are directed toward the striola. In the saccular macula, the kinocilia point away from the striola. Note that, given the utricle and sacculus on both sides of the body, there is a continuous representation of all directions of body movement.
- Figure 14.5. Forces acting on the head and the resulting displacement of the otolithic membrane of the utricular macula. For each of the positions and accelerations due to translational movements, some set of hair cells will be maximally excited, whereas another set will be maximally inhibited. Note that head tilts produce displacements similar to certain accelerations.
- Figure 14.6. Response of a vestibular nerve axon from an otolith organ (the utricle in this example). (A) The stimulus (top) is a change that causes the head to tilt. The histogram shows the neuron's response to tilting in one direction. (B) A response of the same fiber to tilting in the opposite direction. (After Goldberg and Fernandez, 1976.) How Otolith Neurons Sense Linear Forces The structure of the otolith organs enables them to sense both static displacements, as would be caused by tilting the head, and transient displacements caused by translational movements of the head. The mass of the otolithic membrane relative to the surrounding endolymph, as well as the otolithic membrane's physical uncoupling from the underlying macula, means that hair bundle displacement will occur transiently in response to linear accelerations and tonically in response to tilting of the head. Therefore, both tonic and transient information can be conveyed by these sense organs. Figure 14.5 illustrates some of the forces produced by head tilt and linear accelerations on the utricular macula. These properties of hair cells are reflected in the responses of the vestibular nerve fibers that innervate the otolith organs. These vestibular nerve fibers have a steady and relatively high firing rate when the head is upright. The change in firing rate in response to a given movement can be either sustained or transient, decaying back to a tonic level, thereby signaling either absolute head position or linear acceleration. An example of the sustained response of a vestibular nerve fiber innervating the utricle is shown in Figure 14.6 . These responses are from axons in a monkey that was seated in a chair that could be tilted for several seconds to produce a steady force. It is important to note that prior to the tilt, the axon has a high firing rate, which increases or decreases depending on the direction of the tilt. Notice also that the response remains at a high level as long as the tilting force remains constant; thus, such neurons faithfully encode the static force being applied to the head ( Figure 14.6A ). When the head is returned to the original position, the firing level of the neurons returns to baseline value. Conversely, when the tilt is in the opposite direction, the neurons respond by decreasing their firing rate below the resting level ( Figure 14.6B ) and remain depressed as long as the static force continues. In a similar fashion, transient increases or decreass in firing rate from spontaneous levels signal the direction of linear accelerations of the head. In summary, the otolith organs detect linear forces acting on the head, whether by static displacement of hair bundles due to gravity or by transient displacement of hair bundles due to linear accelerations, and this information is preserved in the firing rate of vestibular axons. The range of orientations of hair bundles within the otolith organs enables them to transmit information about linear forces in every direction the body moves (see Figure 14.4C ). The utricle, which is primarily concerned with motion in the horizontal plane, and the sacculus, which is concerned with vertical motion, combine to effectively gauge in three dimensions the linear forces acting on the head at any instant. Tilts of the head off the horizontal plane and translational movements of the head in any direction stimulate a distinct subset of hair cells in the saccular and utricular maculae, while simultaneously suppressing the responses of other hair cells in these organs. Ultimately, variations in hair cell polarity within the otolith organs produce patterns of vestibular nerve fibers activity that, at a population level, can unambiguously encode head position and the forces acting to influence it
- A Primer on (Vestibular) Navigation The function of the vestibular system can be simplified by remembering some basic terminology of classical mechanics. All bodies moving in a three-dimensional framework have six degrees of freedom: three of these are translational and three are rotational. The translational elements refer to linear movements in the x , y , and z axes (the horizontal and vertical planes). Translational motion in these planes (linear acceleration and static displacement of the head) is the primary concern of the otolith organs. The three degrees of rotational freedom refer to a body's rotation relative to the x , y , and z axes and are commonly referred to as roll, pitch, and yaw. The semicircular canals are primarily responsible for sensing rotational accelerations around these three axes.
- Figure 40-6 The bilateral symmetry of the semicircular canals. The horizontal canals on both sides lie in the same plane and therefore are functional pairs. In contrast, the anterior vertical canal on one side and the posterior vertical canal on the opposite side lie in the same plane and are therefore functional pairs. When endolymph begins to move as the result of an acceleration, this fluid presses against one surface of the cupula. Because of its flexibility the cupula bows; the margin into which the hair bundles insert also flexes, thus stimulating the associated hair cells. Because all the hair bundles in each semicircular canal share a common orientation, angular acceleration in one direction depolarizes hair cells and excites afferent axons, while acceleration in the opposite direction hyperpolarizes the receptor cells and diminishes spontaneous neural activity. As with the other receptor organs of the internal ear, the magnitude of the response of the hair cells, as well as that of the afferent axons, is graded with the amplitude of stimulation. In each labyrinth the three canals are almost precisely perpendicular to one another, so that the canals represent accelerations about three mutually orthogonal axes (Figure 40-6). The planes in which the semicircular canals lie do not, however, correspond with the head's major anatomical planes. As its name indicates, the horizontal semicircular canal of each ear lies nearly horizontally with the head in its ordinary, upright position. This canal is accordingly sensitive to rotations about a vertical axis, for example to twisting the neck. The plane in which each anterior vertical semicircular canal lies is slanted about 45° with respect to the coronal plane, so that the lateral extreme of each canal lies rostrally to the medial edge. The planes of the two posterior vertical canals are canted approximately 45° in the opposite direction.
- Figure 14.7. The ampulla of the posterior semicircular canal showing the crista, hair bundles, and cupula. The cupula is distorted by the fluid in the membranous canal when the head rotates. Figure 14.8. Functional organization of the semicircular canals. (A) The position of the cupula without angular acceleration. (B) Distortion of the cupula during angular acceleration. When the head is rotated in the plane of the canal (arrow outside canal), the inertia of the endolymph creates a force (arrow inside the canal) that displaces the cupula. (C) Arrangement of the canals in pairs. The two horizontal canals form a pair; the right anterior canal (AC) and the left posterior canal (PC) form a pair; the left AC and the right PC form a pair. The Semicircular Canals Whereas the otolith organs are primarily concerned with translational movements, the semicircular canals sense head rotations, arising either from self-induced movements or from angular accelerations of the head imparted by external forces. Each of the three semicircular canals has at its base a bulbous expansion called the ampulla ( Figure 14.7 ), which houses the sensory epithelium, or crista , that contains the hair cells. The structure of the canals suggests how they detect the angular accelerations that arise through rotation of the head. The hair bundles extend out of the crista into a gelatinous mass, the cupula , that bridges the width of the ampulla, forming a fluid barrier through which endolymph cannot circulate. As a result, the compliant cupula is distorted by movements of the endolymphatic fluid. When the head turns in the plane of one of the semicircular canals, the inertia of the endolymph produces a force across the cupula, distending it away from the direction of head movement and causing a displacement of the hair bundles within the crista ( Figure 14.8A , B ). In contrast, linear accelerations of the head produce equal forces on the two sides of the cupula, so the hair bundles are not displaced. Unlike the saccular and utricular maculae, all of the hair cells in the crista within each semicircular canal are organized with their kinocilia pointing in the same direction (see Figure 14.2C ). Thus, when the cupula moves in the appropriate direction, the entire population of hair cells is depolarized and activity in all of the innervating axons increases. When the cupula moves in the opposite direction, the population is hyperpolarized and neuronal activity decreases. Deflections orthogonal to the excitatory-inhibitory direction produce little or no response. Each semicircular canal works in concert with a partner located on the other side of the head, which has its hair cells aligned oppositely. There are three such pairs: the two pairs of horizontal canals, and the superior canal on each side working with the posterior canal on the other side ( Figure 14.8C ). Head rotation deforms the cupula in opposing directions for the two partners, resulting in opposite changes in their firing rates ( Box C ). For example, the orientation of the horizontal canals makes them selectively sensitive to rotation in the horizontal plane. More specifically, the hair cells in the canal towards which the head is turning are depolarized, while those on the other side are hyperpolarized. For example, when the head turns to the left, the cupula is pushed toward the kinocilium in the left horizontal canal, and the firing rate of the relevant axons in the left vestibular nerve increases. In contrast, the cupula in the right horizontal canal is pushed away from the kinocilium, with a concomitant decrease in the firing rate of the related neurons. If the head movement is to the right, the result is just the opposite. This push-pull arrangement operates for all three pairs of canals; the pair whose activity is modulated is in the rotational plane, and the member of the pair whose activity is increased is on the side toward which the head is turning. The net result is a system that provides information about the rotation of the head in any direction. The Semicircular Canals Detect Angular Accelerations Angular acceleration occurs whenever an object alters its rate of rotation about an axis. Our head therefore undergoes angular acceleration during turning or tilting motions of the head, rotatory body movements, and turning movements during active or passive locomotion. The three semicircular canals of each vestibular labyrinth detect these angular accelerations and report their magnitudes and orientations to the brain. The name of the semicircular canal aptly reflects its gross structure, which is a roughly semicircular tube of membranous labyrinth extending from the utricle (Figure 40-1). The term “canal” is misleading, however, as the organ is actually a closed tube, nearly 8 mm in overall diameter, and filled with endolymph. Like the otolithic organs, the semicircular canals detect accelerations by means of the inertia of their internal contents. Here, however, it is the mass of endolymph itself that responds to accelerations. Consider the simplest instance of a smoothly increasing rotatory motion, and hence a constant angular acceleration, about an axis passing perpendicularly through the center of a semicircular canal. As the head rotates faster and faster it carries the bony and membranous labyrinths with it. Because of its inertia, however, the endolymph tends to lag behind and therefore rotates within the semicircular canal in a direction opposite that of the head. A cup of coffee can demonstrate the motion of endolymph in a semicircular canal. While gently twisting the cup about its vertical axis, one can observe a particular bubble near the fluid's outer boundary. As the cup begins to turn, the coffee tends to maintain its original orientation in space and thus counterrotates in the vessel. At the conclusion of the turning motion, when the cup decelerates, the coffee moves in the opposite direction. Fluid cannot freely move around the whole of a semicircular canal. Instead, the endolymphatic space of each canal is interrupted by a gelatinous diaphragm, the cupula , that extends across the canal in its widest region, a dilatation termed the ampulla (Figure 40-5). Around most of its perimeter the cupula is attached to the epithelium lining the canal. The portion of the cupula contacting the ampullary crista, however, is less firmly anchored; there the cupula is penetrated by hair bundles extending from a patch of nearly 7000 hair cells.
- Figure 40-7 This view of the horizontal semicircular canals from above shows how the paired canals work together to signal head movement. Because of inertia, rotation of the head in a counterclockwise direction causes endolymph to move clockwise with respect to the canals. This reflects the stereocilia in the left canal in the excitatory direction, thereby exciting the afferent fibers on this side. In the right canal the hair cells are hyperpolarized and afferent firing there decreases. The vestibular labyrinths on the two sides of the head are systematically arranged with respect to one another. The two horizontal canals thus lie in a common plane and hence function together (Figure 40-7). Each anterior vertical canal, in contrast, lies in the same plane as the contralateral posterior vertical canal. Most Movements Elicit Complex Patterns of Vestibular Stimulation Although the actions of the vestibular organs may be separated conceptually and experimentally, as may be the operation of the right and left vestibular labyrinths, actual human movements generally elicit a complex pattern of excitation and inhibition in several receptor organs on both sides of the body. Consider, for example, the act of rising from the driver's seat of an automobile. As one begins to swivel toward the door, both horizontal semicircular canals are strongly stimulated. The simultaneous lateral movement out the car's door stimulates hair cells in both utricles in a pattern that changes continuously as the orientation of the turning head changes with respect to the direction of bodily movement. An appropriately oriented complement of hair cells in each of the saccules is excited, and an oppositely oriented group inhibited, by the vertical acceleration that accompanies rising to a standing position. Finally, the maneuver's conclusion involves linear and angular accelerations opposite to those at the inception. The coffee cup example again confirms the complex pattern of accelerations involved in even a simple movement. One may, for example, examine the result of extending the cup from a position immediately in front of the body to one laterally and at arm's length—the movements involved in serving another person. The angular component of acceleration causes the coffee to rotate within the cup, while the linear component causes the liquid to slosh toward the cup's rim. The conclusion of the movement evokes contrary fluid motions that reflect linear and angular accelerations in the opposite direction. In view of the complexity of the sensory stimuli associated with seemingly simple everyday acts, one may better appreciate why infants need many months of training to support bipedal locomotion. Even as adults we must work diligently to incorporate into reflexes the new patterns of vestibular stimulation associated with new experiences, for example, piloting an airplane. It also seems likely that the need for continual practice by athletes results from constant fine-tuning of vestibular pathways and the associated motor outflows. The complementary and redundant pattern of stimulation of various receptor organs, both within one vestibular labyrinth and between the two internal ears, explains why lesions of the vestibular receptors and pathways can cause disorientation and vertigo. The central nervous system associates a particular pattern of neuronal activity with each motor action in our repertory of behavior. If a component of the vestibular system is excessively active or abnormally silent, the brain receives inappropriate information on acceleration and the reflexes driven by vestibular inputs understandably falter. Only then do we become consciously aware of the vestibular system at work. In the most severe cases a diseased labyrinth must be surgically destroyed in order to relieve the brain of erratic and disabling vestibular signals.
- The ampulla is responsible for the change in rotational movement , as continuous rotation does not stimulate the ampulla. when the head starts moving in a rotationally the endolymph in the semicircular ducts move in the direction opposite to the body’s direction deforming the crista in the duct causes depolarization If the body continues to rotate at a constant rate The endolymph moves at the same direction and speed as the body and stop the movement of hair cells When we suddenly stop moving, the endolymph keeps on moving in the opposite direction hyperpolarization of the hair cells that will tell the brain that we have stopped movement.
- Figure 4: Vestibular nuclei (VN) receive inputs from multiple brain areas. A. Vestibular inputs to the four major vestibular nuclei. B. In addition to inputs from the vestibular receptors, vestibular nuclei receive inputs from other cortical areas, brainstem, cerebellum, and the spinal cord. SVN, LVN, MVN, and IVN are the superior, lateral, medial, and inferior vestibular nuclei, respectively SECOND ORDER VESTIBULAR NEURONS [ edit ] The four main nuclei of the vestibular complex The vestibular complex consist of four main nuclei ((Fig. 4 A)), the medial vestibular nucleus (MVN), the superior vestibular nucleus (SVN), the lateral vestibular nucleus (LVN), and the inferior (or descending) vestibular nucleus (IVN), as well as some other minor subgroups. While there is no real segregation of inputs from afferents, the MVN and SVN receive inputs mostly from horizontal and vertical semicircular canals, respectively. The utricular afferents terminate mainly in the IVN, and also send projections to the LVN,MVN, and SVN. Saccule fibers mainly innervate the LVN and IVN. In addition to these direct projections from vestibular afferents, neurons in the vestibular nuclei also receive inputs from cortical, cerebellar , and other brainstem structures. The impressive convergence of information at the level of the vestibular nuclei is shown in Fig. 4 B. In alert animals these inputs relay somatosensory and visual inputs as well as signals related to eye movements and premotor head movement commands to the vestibular nuclei. As a result, in every day life, these extra-vestibular inputs modify the processing of vestibular information at an early stage of sensory processing.
- THE EFFERENT VESTIBULAR SYSTEM: ANATOMY AND FUNCTION Vestibular efferent neurons are located near the abducens nucleus in the brainstem (Fig. 8 A), Figure 8: Vestibular efferent pathway. A. Efferent fibers originate from a group of cells near the abducens nucleus in the brainstem and project bilaterally to the vestibular periphery. B. Vestibular efferents fibers contact both irregular and regular afferent fibers, as well as type II haircells of the peripheral vestibular system. SVN and LVN are superior and lateral vestibular nuclei, VI represents the abducens nucleus. receive inputs from vestibular afferents and VN, and project back (Fig. 8 B) to vestibular receptors (type II) and afferents (type I and II) (Gacek and Lyon 1974; Goldberg and Fernandez 1980; Rasmussen and Gacek 1958). Vestibular efferent projections are excitatory and increase the resting discharge of the afferents, while decreasing their sensitivity (squirrel monkeys: (Goldberg and Fernandez 1980), toadfish: (Highstein and Baker 1985)). Thus, theoretically, efferents can increase the dynamic range of afferent responses. Furthermore, in frog (Bricout-Berthout et al. 1984; Caston and Bricout-Berthout 1984) and toadfish (Boyle and Highstein 1990; Highstein and Baker 1985) vestibular afferents respond to somatosensory, auditory, or visual stimuli, presumably as a result of inputs mediated by the vestibular efferent system. However, the situation is different in alert primates under normal conditions, since afferent fibers do not carry any extravestibular signals (visual: (Keller 1976), neck proprioception: (Sadeghi et al. 2007b), efference copy of neck motor command: (Cullen and Minor 2002). Nevertheless, efferent-mediated responses are observed in primate afferents in response to high velocity rotations (Sadeghi, Minor, Goldberg, and Cullen, unpublished). Furthermore, vestibular afferents do not play a role in vestibular adaptation (Miles and Braitman 1980) or long-term compensation (Sadeghi et al. 2007b). Thus, the functional role of the efferent system in alert primates has remained elusive.
- Figure 14.9. Response of a vestibular nerve axon from the semicircular canal to angular acceleration. The stimulus (top) is a rotation that first accelerates, then maintains constant velocity, and then decelerates the head. The axon increases its firing above resting level in response to the acceleration, returns to resting level during constant velocity, then decreases its firing rate below resting level during deceleration; these changes in firing rate reflect inertial effects on the displacement of the cupula. (After Goldberg and Fernandez, 1971.) Central Vestibular Pathways: Eye, Head, and Body Reflexes Figure 14.10. Connections underlying the vestibulo-ocular reflex. Projections of the vestibular nucleus to the nuclei of cranial nerves III (oculomotor) and VI (abducens). The connections to the oculomotor nucleus and to the contralateral abducens nucleus are excitatory (red), whereas the connections to ipsilateral abducens nucleus are inhibitory (black). There are connections from the oculomotor nucleus to the medial rectus of the left eye and from the adbucens nucleus to the lateral rectus of the right eye. This circuit moves the eyes to the right, that is, in the direction away from the left horizontal canal, when the head rotates to the left. Turning to the right, which causes increased activity in the right horizontal canal, has the opposite effect on eye movements. The projections from the right vestibular nucleus are omitted for clarity. The vestibular end organs communicate via the vestibular branch of the eighth cranial nerve with targets in the brainstem and the cerebellum that perform much of the processing necessary to compute head position and motion. As with the cochlear nerve, the vestibular nerves arise from a population of bipolar neurons, the cell bodies of which in this instance reside in the vestibular nerve ganglion (also called Scarpa's ganglion ; see Figure 14.1 ). The distal processes of these cells innervate the semicircular canals and the otolith organs, while the central processes project via the vestibular portion of cranial nerve VIII to the vestibular nuclei (and also directly to the cerebellum) ( Figure 14.10 ). Because vestibular and auditory fibers run together in the eighth nerve, damage to this structure often results in both auditory and vestibular disturbances. The vestibular nuclei are important centers of integration, receiving input from the vestibular nuclei of the opposite side, as well as from the cerebellum and the visual and somatic sensory systems. One of the main functions of the vestibular system is to coordinate head and eye movements (other functions include protective or escape reactions; see Box D ). The vestibulo -ocular reflex ( VOR ) in particular is a mechanism for producing eye movements that counter head movements, thus permitting the gaze to remain fixed on a particular point ( Box C ; see also Chapter 20 ). For example, activity in the left horizontal canal induced by leftward rotation of the head excites neurons in the left vestibular nucleus and results in reflexive eye movements to the right. This effect is due to excitatory projections from the vestibular nucleus to the contralateral nucleus abducens that, along with the oculomotor nucleus, help execute conjugate eye movements. For instance, horizontal movement of the two eyes toward the right requires contraction of the left medial and right lateral rectus muscles. Vestibular nerve fibers originating in the left horizontal semicircular canal project to the medial and lateral vestibular nuclei (see Figure 14.10 ). Excitatory fibers from the medial vestibular nucleus cross to the contralateral abducens nucleus, which has two outputs. One of these is a motor pathway that causes the lateral rectus of the right eye to contract; the other is an excitatory projection that crosses the midline and ascends via the medial longitudinal fasciculus to the left oculomotor nucleus, where it activates neurons that cause the medial rectus of the left eye to contract. Finally, inhibitory neurons project from the medial vestibular nucleus to the left abducens nucleus, directly causing the motor drive on the lateral rectus of the left eye to decrease and also indirectly causing the right medial rectus to relax. The consequence of these several connections is that excitatory input from the horizontal canal on one side produces eye movements toward the opposite side. Therefore, turning the head to the left causes eye movements to the right. In a similar fashion, head turns in other planes activate other semicircular canals, causing other appropriate compensatory eye movements. The rostro-caudal set of cranial nerve nuclei involved in the VOR (i.e., the vestibular, abducens and oculomotor nuclei), as well as the VOR's persistence in the unconscious state, make this reflex especially useful for detecting brainstem damage in the comatose patient (see Box C ). Loss of the VOR can have severe consequences. A patient with vestibular damage finds it difficult or impossible to fixate on visual targets while the head is moving, a condition called oscillopsia . If the damage is unilateral, the patient usually recovers the ability to fixate objects during head movements. However, a patient with bilateral loss of vestibular function has the persistent and disturbing sense that the world is moving when the head moves. The underlying problem in such cases is that information about head and body movements normally generated by the vestibular organs is not available to the oculomotor centers, so that corrective eye movements cannot be made. Descending projections from the vestibular nuclei are essential for postural adjustments of the head and body. As with the VOR, these postural reflexes are extremely fast, in part due to the small number of synapses interposed between the vestibular organ and the relevant motor neurons ( Box D ). Axons from the medial vestibular nucleus descend in the medial longitudinal fasciculus to reach the upper cervical levels of the spinal cord ( Figure 14.11 ). This pathway regulates head position by reflex activity of neck muscles in response to stimulation of the semicircular canals from rotational accelerations of the head. For example, during a downward pitch of the body (e.g., tripping), the superior canals are activated and the head muscles reflexively pull the head up. The dorsal flexion of the head initiates other reflexes, such as forelimb extension and hindlimb flexion, to stabilize the body and protect against a fall (see Chapter 17 ). The inputs from the otolith organs project mainly to the lateral vestibular nucleus, which in turn sends axons in the lateral vestibulospinal tract to the spinal cord (see Figure 14.11 ). The input from this tract exerts a powerful excitatory influence on the extensor (antigravity) muscles. When hair cells in the otolith organs are activated, signals reach the medial part of the ventral horn. By activating the ipsilateral pool of motor neurons innervating extensor muscles in the trunk and limbs, this pathway mediates balance and the maintenance of upright posture. Decerebrate rigidity, which is characterized by rigid extension of the limbs, arises when the brainstem is transected above the level of the vestibular nucleus. The tonic activation of extensor muscles in this instance suggests that the vestibulospinal pathway is normally strongly suppressed by descending projections from higher levels of the brain, especially the cerebral cortex (see also Chapter 17 ) How Semicircular Canal Neurons Sense Angular Accelerations Like those that innervate the otolith organs, the vestibular fibers originating from bipolar neurons in Scarpa's ganglion that innervate the semicircular canals exhibit a high and steady spontaneous firing rate. As a result, they can transmit information by either increasing or decreasing their firing rate. As already suggested, such bidirectional responses enable the afferent nerves to faithfully follow the receptor potentials, and thus to more effectively encode head movements. The bidirectional responses of fibers innervating the hair cells of the semicircular canal have been studied by recording the axonal firing rates in a monkey's vestibular nerve. Seated in a chair that could be rotated, the monkey was first rotated at an accelerated rate, then at constant velocity for several seconds, and finally the chair was decelerated to a stop ( Figure 14.9 ). The maximum firing rates observed correspond to the period of acceleration; the maximum inhibition corresponds to the period of deceleration. During the constant-velocity phase, the response adapts so that the firing rate subsides to resting level; after the movement stops, the neuronal activity decreases transiently before returning to the resting level. Neurons innervating paired canals have a complimentary response pattern. Note that the rate of adaptation (on the order of tens of seconds) corresponds to the time it takes the cupula to return to its undistorted state (and for the hair bundles to return to their undeflected position); adaption therefore can occur even while the head is still turning, as long as a constant angular velocity is maintained. Such constant forces are rare in nature, although they are encountered on ships, airplanes, and space vehicles, where prolonged acceleratory arcs are sometimes described.
- Figure 14.11. Descending projections from the medial and lateral vestibular nuclei to the spinal cord. The medial vestibular nuclei project bilaterally in the medial longitudinal fasciculus to reach the medial part of the ventral horns and mediate head reflexes in response to activation of semicircular canals. The lateral vestibular nucleus sends axons via the lateral vestibular tract to contact anterior horn cells innervating the axial and proximal limb muscles. Neurons in the lateral vestibular nucleus receive input from the cerebellum, allowing the cerebellum to influence posture and equilibrium.
- Figure 14.12. Thalamocortical pathways carrying vestibular information. The lateral and superior vestibular nuclei project to the thalamus. From the thalamus, the vestibular neurons project to the vicinity of the central sulcus near the face representation. Sensory inputs from the muscles and skin also converge on thalamic neurons receiving vestibular input (see Chapter 9 ). Vestibular Pathways to the Thalamus and Cortex In addition to these several descending projections, the superior and lateral vestibular nuclei send axons to the ventral posterior nuclear complex of the thalamus, which projects to two cortical areas relevant to vestibular sensations ( Figure 14.12 ). One cortical target is just posterior to the primary somatosensory cortex, near the representation of the face; the other is at the transition between the somatic sensory cortex and the motor cortex (Brodmann's area 3a; see Chapter 9 ). Electrophysiological studies of individual neurons in these areas show that the relevant cells respond to proprioceptive and visual stimuli as well as to vestibular stimuli. Many of these neurons are activated by moving visual stimuli as well as by rotation of the body (even with the eyes closed), suggesting that these cortical regions are involved in the perception of body orientation in extrapersonal space HIGHER ORDER VESTIBULAR PROCESSING [edit] Vestibulo-cerebellum There are five main regions of the cerebellum that receive either primary (i.e. from afferents) or secondary (i.e. from vestibular nuclei) vestibular input including: i) the nodulus and ventral uvula, ii) the flocculus and ventral paraflocculus, iii) the oculomotor vermis of posterior lobe, iv) lobules I-V of the anterior lobe, and v) the deep cerebellar nuclei. As described below, each of these regions makes an important contribution to the processing of vestibular sensory information. i) The vestibular nuclei are reciprocally interconnected with the nodulus/uvula of the cerebellum (Wearne et al. 1998). These areas of the cerebellum make significant contributions to the computation of inertial motion (Angelaki and Hess 1995; Wearne et al. 1998). ii) The flocculus and adjoining paraflocculus is involved in the generation and the plasticity of compensatory eye movements, including visual ocular following reflexes (i.e. smooth pursuit and the optokinetic reflex) and the VOR (Buttner and Waespe 1984; Lisberger and Fuchs 1978; Lisberger et al. 1994a; Lisberger et al. 1994b; Miles et al. 1980a; Miles et al. 1980b; Noda and Suzuki 1979). iii) Neurons in lobules VI and VII of the vermis, which is called the oculomotor vermis, contribute to visual-vestibular processing (Sato and Noda 1992; Suzuki and Keller 1988, 1982). In addition to its vestibular inputs, this cerebellar region receives eye movement signals from the nucleus preopositus (Belknap and McCrea 1988) as well as pursuit-related inputs from the dorsolateral pontine nuclei (Brodal 1979; Yamada and Noda 1987) - a region to which cortical pursuit areas MT and MST both project (Glickstein et al. 1980). iv) The anterior region of the cerebellar vermis (lobules I-V) encodes both vestibular and neck proprioceptive-related signals (Manzoni et al. 2004; Manzoni et al. 1998a, 1998b; Manzoni et al. 1999) and is thought to control of vestibulo-spinal reflexes. The integration of vestibular and proprioceptive information ensures that the motor responses produced by these reflexes are appropriate to maintain body stability. v) The signal processing done in the fastigial nucleus of the deep cerebellar nuclei is tightly linked to the vestibular system. It receives both primary and secondary vestibular inputs as well as input from the cerebellar vermis. The fastigial nucleus plays an important role in the generation of postural reflexes and orienting behaviors, and accordingly projects to brainstem structures that control these behaviors including the vestibular nuclei and medial reticular formation. Many neurons in this area intrgarate vestibular and propiciocpetive inputs, and in turn, encode vestibular signals in a body-centered reference frame (Kleine et al. 2004; Shaikh et al. 2004 Summary The receptor cells of the vestibular system are located in the otolith organs and the semicircular canals of the inner ear and provide information about the motion and position of the body in space. The otolith organs provide information necessary for postural adjustments of the somatic musculature, particularly the axial musculature, when the head tilts in various directions or undergoes linear accelerations. This information represents linear forces acting on the head that arise through static effects of gravity or from translational movements. The semicircular canals, in contrast, provide information about rotational accelerations of the head. This latter information generates reflex movements that adjust the eyes, head, and body during motor activities. Among the best studied of these reflexes are eye movements that compensate for head movements, thereby stabilizing the visual scene when the head turns. Input from all the vestibular organs is integrated with input from the visual and somatic sensory systems to provide perceptions of body position and orientation in extrapersonal space
- WHEN WE MOVE WE ARE usually unaware of the complex neuromuscular processes that control our posture. But postural control is obvious enough when we accidentally fall or when disease damages parts of the postural system. The mechanical problem of maintaining posture is particularly challenging for erect bipeds. Even with the more stable four-legged animals successful postural control is no mean achievement. Imagine a predator chasing its prey at full gallop and then suddenly changing direction. Before doing so it must first throw its center of gravity into the curve, correctly predicting the new gravito-inertial force vector introduced by centrifugal force. Humans also make this kind of postural adjustment, usually without thinking. In fact, conscious control can be disastrous. One might think that to make a rightward turn on a bicycle the rider simply rotates the handlebars to the right. Wrong! Rotation to the right would move the center of gravity to the left, and the cyclist would quickly crash to the ground. Instead, without realizing it, the cyclist first turns the handlebars briefly leftward to generate a centrifugal force that moves the body rightward into the intended curve. Only then does he steer the bicycle to the right until the new force vector properly lines up with wheel contact on the ground. Children must learn this postural adjustment when switching from a tricycle (in which the handlebars must first be turned in the direction the rider wants to go) to a bicycle. The postural system must therefore meet three main challenges. It must maintain a steady stance (balance) in the presence of gravity, it must generate responses that anticipate volitional goal-directed movements, and it must be adaptive. Posture and Equilibrium How should posture be defined? What are its behavioral goals? Posture is essentially the relative position of the various parts of the body with respect to one another (the egocentric coordinate system ) and to the environment (the exocentric coordinate system ). A third frame of reference is that of the gravitational field (the geocentric coordinate system ). The orientation of a body part can be described in terms of each of these frameworks, depending on the behavioral context. For example, knowing the position of the head relative to the environment is important in stabilizing vision, while knowing its position relative to the rest of the body is important in maintaining erect posture. Regulation of posture with respect to gravity is obviously important in maintaining postural equilibrium , which may be defined as the state in which all forces acting on the body are balanced so that the body rests in an intended position ( static equilibrium ) or is able to progress through an intended movement without losing balance ( dynamic equilibrium ). Postural control may have different goals under different circumstances, such as longitudinal alignment of the whole body to maintain a steady, erect stance; remodeling of stance in preparation for a voluntary movement; shaping of the body for display purposes, as in dance; maintenance of balance, as on the gymnast's beam; or conservation of energy in a demanding task. Different animals may use quite different postural strategies to achieve a steady stance in the face of gravity. Thus, as a result of size-dependent mechanics ( allometry ), the elephant mechanically locks its legs in line with the gravity vector to minimize the muscle forces supporting its heavy body. In contrast, small quadrupeds tense the muscles around flexed limb joints, thus permitting rapid responses to danger. (Tensing the muscles around flexed joints enhances muscle stiffness; since stiff muscle pairs mechanically resist change virtually instantaneously and hence well ahead of any active reflex response, opposition to unexpected perturbations is optimal.) Posture and Movement Both of these strategies are used by humans. We stand like elephants. Instead of flexing the knees as if to spring forward, we lock the knee joints so that the legs become static and structural, thus off-loading antigravity muscles until they are needed for movement. On the other hand, like smaller animals we flex the weight-supporting limbs in preparation for intended movement, as at the start of a sprint.
- Figure 41-1 Postural adjustments require anticipatory motor actions. (Adapted from Lee et al. 1995.) A. Rear view of a subject standing on a force-transducing platform and lifting the right leg sideways to the right. The numbered points identify points on the body from which the image in part B is constructed. B. A record of trunk and limb movements during this maneuver. The black lines represent the subject with both feet resting symmetrically on the ground. The colored lines, separated by 50 ms intervals and moving in the direction of the arrows, show the moment-to-moment pattern of postural adjustment employed to reestablish static equilibrium on the left leg without falling. C. Kinematic records show the hip and the shoulders beginning to move long before the ankle starts to rise. The center of pressure registered by the force-transducing platform first moves outward toward the leg that is to be lifted in order to move the center of gravity over the supporting leg. Then, as the shoulders and the hip begin to move, it moves back under the supporting leg to maintain the new posture. Postural Readjustment Must Be Preceded by Anticipatory Motor Action Now consider what happens when one leg is actively lifted sideways while a subject maintains balance on the other. Since the center of gravity's force vector is initially projected midway between the two feet, the body would fall to the side of the lifted leg if no anticipatory action were taken. This is readily demonstrated when one stands sideways against a solid wall and then raises the outside leg. It cannot be done without falling away from the wall! To maintain balance, the voluntary movement must be preceded by a counterbalancing movement that shifts the center of gravity over the leg on which the person intends to remain standing. This apparently simple procedure invokes a complex set of interactive responses, however. Some 600 ms before the rising ankle begins to move, a subject will briefly move the center of pressure (recorded on a platform) toward the outside foot so as to provide the necessary force for transferring the body's center of mass over the inside foot. Since shoulder and hip movements are driven by the changing center of pressure, they too are initiated before the ankle begins to move. As the shoulders and the hips begin to move, the center of pressure returns to the support leg (Figure 41-1).
- Figure 17.4. Anticipatory maintenance of body posture. At the onset of a tone, the subject pulls on a handle, contracting the biceps muscle. To ensure postural stability, contraction of the gastrocnemius muscle precedes that of the biceps. Figure 17.5. Feedforward and feedback mechanisms of postural control. Feedforward postural responses are “preprogrammed” and typically precede the onset of limb movement (see Figure 17.4 ). Feedback responses are initiated by sensory inputs that detect postural instability. The way the upper motor neurons of the reticular formation maintain posture can be appreciated by analyzing their activity during voluntary movements. Even the simplest movements are accompanied by the activation of muscles that at first glance seem to have little to do with the primary purpose of the movement. For example, Figure 17.4 shows the pattern of muscle activity that occurs as a subject uses his arm to pull on a handle in response to an auditory tone. Activity in the biceps muscle begins about 200 ms after the tone. However, as the records show, the contraction of the biceps is accompanied by a significant increase in the activity of a proximal leg muscle, the gastrocnemius (as well as many other muscles not monitored in the experiment). In fact, contraction of the gastrocnemius muscle begins well before contraction of the biceps. These observations show that postural control entails an anticipatory, or feedforward, mechanism (Figure 17.5). As part of the motor plan for moving the arm, the effect of the impending movement on body stability is “evaluated” and used to generate a change in the activity of the gastrocnemius muscle. This change actually precedes and provides postural support for the movement of the arm. In the example given here, contraction of the biceps would tend to pull the entire body forward, an action that is opposed by the contraction of the gastrocnemius muscle. In short, this feedforward mechanism “predicts” the resulting disturbance in body stability and generates an appropriate stabilizing response. The importance of the reticular formation for feedforward mechanisms of postural control has been explored in more detail in cats trained to use a forepaw to strike an object. As expected, the forepaw movement is accompanied by feedforward postural adjustments in the other legs to maintain the animal upright. These adjustments shift the animal's weight from an even distribution over all four feet to a diagonal pattern, in which the weight is carried mostly by the contralateral, nonreaching forelimb and the ipsilateral hindlimb. Lifting of the forepaw and postural adjustments in the other limbs can also be induced in an alert cat by electrical stimulation of the motor cortex. After pharmacological inactivation of the reticular formation, however, electrical stimulation of the motorcortex evokes only the forepaw movement, without the feedforward postural adjustments that normally accompany them.
- Figure 41-2 Anticipatory motor action in response to postural disturbance adapts to the behavioral context. The illustrations show the postural activity of the gastrocnemius muscle in four behavioral contexts. (Adapted from Cordo and Nashner 1982.) A. The subject stands on a firm platform and pulls on a fixed handle as soon as possible after an auditory cue. To maintain posture, backward-acting contraction of the leg muscle (gastrocnemius) starts before the biceps begin pulling the handle. B. When the chest is supported and the handle is suddenly pulled forward, there is a very early reflex response in the biceps and the gastrocnemius remains silent. C. When the handle suddenly pulls the unsupported subject forward, the early biceps reflex is suppressed so that counterbalancing arm and leg muscles can act simultaneously. D. Finally, when the foot support unexpectedly slides forward, tilting the subject backward, the gastrocnemius is silent (otherwise it would tilt the subject further back) and an early biceps response is brought into play. E. The feed-forward and feedback components of postural control. (Adapted from Gahéry and Massion 1981.) Postural Control Can Be Adapted to Suit Specific Behaviors Anticipatory action is remarkably adaptable and varies according to behavioral demand (Figure 41-2). One can easily demonstrate the dependence of postural control on behavioral context, or postural set. First, place the right upper arm beside the body with the forearm horizontal so that the elbow of the arm is at a right angle. Then, while actively maintaining this posture with the eyes closed, try to press that forearm down with the left hand. If one suddenly releases the downward pressure of the left hand, there is little or no movement of the right forearm. Anticipatory suppression of the right forearm flexors is synchronized with the predicted removal of the left hand. Next, again with the eyes shut, have a colleague apply downward pressure on the right forearm, and then ask that it be suddenly removed without warning. Now the right forearm flies upward. Anticipatory responses depend on feed-forward control. The central command for a voluntary limb movement is associated with a simultaneous feed-forward command anticipating an expected postural perturbation. Postural adjustments to unexpected disturbances depend on feedback, however. These adjustments may be relatively simple and fast, as in the biceps stretch reflex (Figure 41-2B), but usually they are the product of complex motor reactions that are learned and released as a whole (Figure 41-2C). The interaction of feed-forward and feedback control of posture is summarized in Figure 41-2E.
- Figure 41-3 Appropriate anticipatory responses to postural disturbances can be learned. (Adapted from Nashner 1976.) A. Backward movement of the sliding platform tilts the body forward, calling for countervailing action in the stretched gastrocnemius to maintain balance. In successive trials the muscular response is enhanced and its latency reduced. B. When the platform is tilted up, action by the gastrocnemius would worsen the backward body tilt. Accordingly, in successive trials the muscle's response is decreased, with a corresponding decrease of backward sway. While the basic elements of postural control are innate, they nevertheless can be greatly modified by learning. The rigid stance of the decerebrate quadruped is innate, as is the stereotyped response of a standing quadruped when it loses the support of one or more limbs. If support from diagonally opposite legs is unexpectedly withdrawn, the decerebrate animal actively reacts by supporting itself on the other pair of legs. The advantage of this strategy is that the center of gravity need not be moved, although it does create a problem with stabilization. If an intact animal is trained to expect loss of support from one paw, it adopts a quite different strategy at the time of unloading: The center of gravity is actively moved within the triangular support of the other three limbs. This action provides support and ensures subsequent postural equilibrium. Russian investigators in the early twentieth century used classical Pavlovian conditioning to show that animals can learn anticipatory feed-forward patterns of response. They trained animals with deafferented limbs to remove the support of one limb by actively lifting one paw in response to an auditory cue. Recordings of postural muscle activity showed that the trained animals always appropriately activated the antigravity muscles of the other three limbs before raising the paw. Later, Lew Nashner devised an ingenious way to demonstrate adaptive learning of postural control in humans. The subject stands erect on a platform, which can be made to slide backward without tilting or to tilt toe-up without sliding. Both maneuvers stretch the gastrocnemius. But while forward sway calls for contraction of the muscle, backward sway calls for its relaxation. In both conditions there is clear evidence that the subject learns the appropriate response (Figure 41-3). While the basic elements of postural control are innate, they nevertheless can be greatly modified by learning. The rigid stance of the decerebrate quadruped is innate, as is the stereotyped response of a standing quadruped when it loses the support of one or more limbs. If support from diagonally opposite legs is unexpectedly withdrawn, the decerebrate animal actively reacts by supporting itself on the other pair of legs. The advantage of this strategy is that the center of gravity need not be moved, although it does create a problem with stabilization. If an intact animal is trained to expect loss of support from one paw, it adopts a quite different strategy at the time of unloading: The center of gravity is actively moved within the triangular support of the other three limbs. This action provides support and ensures subsequent postural equilibrium. Russian investigators in the early twentieth century used classical Pavlovian conditioning to show that animals can learn anticipatory feed-forward patterns of response. They trained animals with deafferented limbs to remove the support of one limb by actively lifting one paw in response to an auditory cue. Recordings of postural muscle activity showed that the trained animals always appropriately activated the antigravity muscles of the other three limbs before raising the paw. Later, Lew Nashner devised an ingenious way to demonstrate adaptive learning of postural control in humans. The subject stands erect on a platform, which can be made to slide backward without tilting or to tilt toe-up without sliding. Both maneuvers stretch the gastrocnemius. But while forward sway calls for contraction of the muscle, backward sway calls for its relaxation. In both conditions there is clear evidence that the subject learns the appropriate response (Figure 41-3).
- Figure 41-5 The adaptability of the postural components of locomotion is demonstrated in the following experiment. The subject first walks in place (ie, remains stationary in space), with the eyes open, on the perimeter of a rotating circular disc for 1-2 hours (A). Throughout this period the space-stationary trunk is associated with a constant relative angular velocity of the stance foot, since the foot rotates with the turntable when planted on it. Before adaptation to the rotating treadmill a blindfolded subject can walk straight ahead on firm ground quite well. After adaptation the blindfolded subject can no longer walk straight ahead. When trying to do so he unknowingly walks in a circular path on the ground by steadily rotating the trunk relative to the stance foot (B), even when the resulting body angular velocity is well above the vestibular threshold of rotational sensation. In contrast, after adaptation the blindfolded subject can readily propel a wheelchair straight ahead with the hands (C), demonstrating that the motor learning is localized in the podokinetic system for sensory-motor control of trunk rotation relative to the stance foot. (Adapted from Gordon et al. 1995.) Adaptive Postural Control Is Learned During Locomotion A normal blindfolded person can walk straight ahead quite well. This task requires precise neuromuscular control of horizontal trunk rotation relative to the stance foot on the ground in order to prevent curvature in the trajectory of locomotion. This neuromuscular control must be learned. In a recent series of experiments subjects were required to walk on the perimeter of a rotating disc without moving in space, much like walking on a treadmill. After an hour or so of this unusual experience, blindfolded subjects could no longer walk straight ahead on firm ground. Instead, they unknowingly walked in curved trajectories, even though the rate of rotation relative to space was then well above the threshold of vestibular sensation. In contrast, the same subjects were able to propel themselves manually in a wheelchair along a straight path, demonstrating that the learning was restricted to the postural components exposed to the adaptive stimulus, namely, those of the lower limb locomotor system (Figure 41-5). That is, the adaptive learning took place within a “bottom-up” flow of postural information. In contrast, control of spatial relations between the head and the trunk by the vestibular-neck postural system involves “top-down” information flow.
- Figure 41-6 The vestibulospinal and cervicospinal reflexes act synergistically to maintain body posture whether the head and trunk move together or independently. A. The head and the trunk are in the normal standing position with both forelegs extended. B. Passive rotation of both the head and the trunk to the left leads to left leg extension and right leg flexion. This pattern of reflex response is a purely vestibulospinal one since there is no independent movement of the head and the trunk. C. Tilting the trunk left-side down while keeping the head stationary in space produces the same behavior as in B. Here, however, the reflex response is purely cervicospinal. D. When the head is rotated relative to both gravity and the trunk, the two reflexes oppose one another and limb extension remains balanced, thus allowing the head to rotate relative to the trunk without postural disturbance. Vestibular and Neck Contributions When subjects are rotated in one direction for a minute or so and then try to stand up while suddenly tilting the head relative to the trunk, they throw themselves to the ground because of powerful but inappropriate vestibulospinal postural reflexes (don't try this without restraint—it's dangerous!). Why are these reflexes so inappropriate in these circumstances? The grossly misleading vestibulospinal response that occurs after prolonged rotation becomes mismatched to the concurrent but normal afferent stimulation of neck afferents introduced by head tilt. In normal circumstances we can sharply rotate the head relative to the trunk without any disturbance of equilibrium, but only because of the precisely calibrated offset of vestibulospinal reflexes by neck afferent signals. With the trunk stationary, the two separate but powerful influences normally cancel one another perfectly so that their presence in the central nervous system goes unnoticed by the subject. When the trunk moves relative to space, the separate roles of the two pathways that project to the spinal cord become apparent. Vestibular and Neck Reflexes Are Subject to Volitional Control Many now-classical behavioral studies done early in the twentieth century identified a series of vestibular-neck (vestibulocollic), vestibulospinal, neck-neck (cervicocollic), and cervicospinal reflexes, all of which appear to be innate. Since the vestibular sensory signal relays a message of head movement relative to inertial space (Chapter 40), it is not surprising that the vestibulocollic reflex plays an important role in stabilizing the head relative to space. Significantly, however, the vestibulocollic reflex acts in a fundamentally different way from the more familiar vestibulo-ocular reflexes that stabilize the eyes (Chapter 39). When the vestibulocollic reflex acts on neck muscles to stabilize the head relative to space, the motor response, by opposing the perturbing head movement, necessarily tends to null the vestibular signal at its source. In other words, this reflex functions as an error-activated negative feedback system In contrast, the cervicocollic reflex responds to neck muscle stretch and joint afferents and therefore stabilizes the head relative to the trunk (rather than to space). Consequently, when the trunk is stationary the vestibulocollic and cervicocollic reflexes collaborate in head stabilization. But when the trunk turns relative to the head, as when one looks at a fixed object while turning a corner, the cervicocollic reflex must be suppressed so that the vestibulocollic reflex alone stabilizes the head. Thus, even though innate, these reflexes can be brought under higher control to suit the intended pattern of voluntary movement. Vestibulospinal and Cervicospinal Reflexes Collaborate in Maintaining Postural Stability The innate vestibulospinal and cervicospinal reflexes automatically maintain postural stability. For example, if the head and trunk of a quadruped are both passively tilted to the left, vestibulospinal responses extend the left limbs and flex the right ones to oppose the perturbation. If the trunk alone is passively tilted left while the head is held stationary, the cervicospinal response produces opposition to the tilt. However, if the head is rotated while the trunk remains stationary, these two reflexes oppose one another, so that the animal's stance is not disturbed (Figure 41-6). Again, even though these reflexes are innate, they can be altered or superseded by higher brain centers to suit the behavioral context. Another innate vestibulospinal response occurs during sudden falls. Humans dropped unexpectedly exhibit a stereotyped extensor response in the antigravity muscles of the lower limbs that serves to break the fall. But the latency of the fixed response is less than 100 ms, so this response is useful only for falls from heights great enough to allow the response to come into play. Here, too, there is a need for flexibility in an innate reflex response, which must be integrated with volitional control based on the subject's assessment of the distance to the ground. When a cat with clear vision is suddenly dropped, the short-latency vestibulospinal response is superimposed upon by context-dependent muscle activity, which is nicely timed within a millisecond or two of the anticipated moment of landing. Spinal stretch reflexes alone would not serve to resist the landing from any height because the latency of such a reaction, which includes the electromechanical coupling time, would be far too long to permit muscular deceleration even by the monosynaptic stretch reflex. This long latency creates a problem when, for example, one descends a step of unanticipated depth. If the time of fall is less than the latency of the vestibulospinal response, muscular deceleration is not possible, and a dangerous skeletal impact is bound to occur.
- Figure 41-7 The vestibular contribution to postural adjustment depends on whether the primary perturbation is to the feet or the head. (Adapted from Horak et al. 1994.) A. Sudden backward movement of the feet leads to a large and early response in the gastrocnemius muscle but no relevant action in the trapezius or sternomastoid neck muscles. Although the head does move when the feet are displaced, the resulting vestibular activity cannot be responsible for the gastrocnemius response since the latency of the response is too brief. In fact, the latency is that of the stretch reflex. B. In contrast, sudden forced acceleration of the head produces the opposite effects: a strong, short-latency response in the neck muscle and little or no response in the gastrocnemius. The vestibular contribution to postural control depends on the kind of perturbation imposed. For example, if the platform on which a subject stands is suddenly and unexpectedly moved backward, patients with defective vestibular labyrinths make normal postural muscle reactions to the forward-induced fall. If the head rather than the foot position is perturbed, the same patients cannot make normal short-latency corrective responses (Figure 41-7). A primary function of the vestibular response is to stabilize the head in space. When a normal subject balances on a small support surface, a so-called hip balancing strategy is brought into play. With the head held steady, one maintains balance by continually shifting the pelvic girdle back and forth. Under conditions in which a normal subject copes quite well, the patient with a labyrinth defect quickly loses balance because the vestibular components of head stabilization are lacking. Furthermore, when cats who have just been labyrinthectomized turn their heads relative to both trunk and space, they immediately lose their balance and fall because the cervicospinal reflexes are no longer balanced by matching vestibular input.
- Figure 41-8 After unilateral removal of a cat's vestibular labyrinth, the ultimate degree of recovery of the animal's ability to maintain dynamic equilibrium is highly dependent on postoperative management. The curves show recovery patterns in the ability to walk along a beam that can be rotated at various speeds. (The curves begin 12 days after surgery because before that time none of the cats could perform the task even on a static beam.) The unrestrained cat achieved full postoperative recovery after about 8 weeks of practice. A period of 1 week's physical restraint after full recovery had no effect (A). Restraint in the middle of the recovery period seriously reduced the otherwise attainable degree of recovery (B). One week of restraint immediately after operation reduced the ultimate level of recovery to only 40% of the potential level (C). (Adapted from Xerri and Lacour 1980.) Perhaps a similar rationale explains why, while a normal cat readily learns to walk along a rotating beam without falling, an animal with bilateral labyrinthectomy never recovers this remarkable ability. In contrast, the unilaterally defective animal can relearn this demanding postural motor task through adaptive mechanisms of the kind discussed in the next section. Significantly, the potential for long-term recovery is highly dependent on postoperative management. Animals normally recover completely within about 6 weeks. But if the animal is artificially constrained for 1 week during the first 10 days after surgical lesioning, the ultimate degree of recovery is severely impaired; no more than about 40% of the possible extent of recovery is achieved. Constraint in the middle of the recovery period severely prejudices the potential achievement, but constraint after full recovery has no effect since the motor learning is then complete (Figure 41-8). Perceptual Correlates The powerful effects of illusory self-motion indicate that cognitive perception plays an important role in postural control. To study this matter Thomas Mergner and his colleagues assembled a device that allows the human head, trunk, and feet to be independently turned relative to space and one another. The subject's perception in the dark of the relative movement of these body parts and their place in space were quantitatively recorded while turning first the whole body relative to space, then the trunk relative to the stationary head, and finally the feet relative to the stationary trunk. As expected, rotation of the whole body relative to space showed the normal high-pass response typical of the vestibular system, with a sensation threshold of around 2°/s and an associated sensation “decay” time constant of 10-15 s. However, perception of the relative movement of the head, trunk, and feet was found to depend on which body parts were moved and the nature of the perceptual task. For example, when the trunk was turned at various sinusoidal frequencies relative to the stationary head, the subject's estimate of trunk rotation relative to space reflected the high-pass dynamics of vestibular sensation, even though rotational vestibular stimulation was completely lacking in this situation. On the other hand, the subject's estimate of head rotation relative to the trunk had no vestibular-like dynamics, and the sensation threshold was decreased by an order of magnitude to around 0.2°/s. Similarly, when the foot and the trunk were turned relative to one another, the perception of foot rotation relative to the trunk reflected vestibular-like high-pass dynamics, while that of trunk rotation relative to the feet reflected the same low-pass characteristics and low threshold as those of head sensation relative to the trunk. Thus there appear to be two discrete modes of perceiving these postural relations. One, stemming from the vestibular system, takes the head as a reference and projects information in a “top-down” manner from head to trunk to feet. The other takes the stable ground as a reference and projects in a “bottom-up” manner from feet to trunk to head. The top-down pathway faithfully registers high-frequency components of movement, such as those that occur during jogging, for which the high-pass filtering of the vestibular system is well adapted. The bottom-up pathway serves the complementary role of registering low-frequency and low-amplitude angular movements, such as those encountered during a change of direction while walking. Our conscious ideas about postural changes have physiological consequences. This is demonstrated by an experiment in which the whole body is passively rotated while the subject tries to visually fixate an imagined stationary target in the dark. If gaze direction drifts away from the target because of underperformance of the compensatory vestibulo-ocular reflex, the intended goal tends nevertheless to be achieved by internally generated corrective saccadic eye movements (termed vestibular memory-contingent saccades ). These corrective oculomotor saccades are presumably activated when there is a difference between a valid vestibular perceptual estimate of rotation relative to the target and a calibrated motor copy of the total angle of compensatory eye movement relative to the head. The corrective saccades occur only if the subject consciously tries to maintain target fixation, however. As observed by the Russian physiologist Viktor Gurfinkel, “The system of internal (perceptual) representation plays a dominant role in postural control.” An Overall View The goal of postural control is to orient body parts relative to one another and the external world without loss of balance. Posture must be controlled both while the body is still (static equilibrium) and during movement (dynamic equilibrium). In the dynamic states of natural behavior voluntary movement can perturb postural equilibrium, but knowledge of these potential perturbations is built into the motor program and used to offset their adverse effects ahead of the event by anticipatory (feed-forward) motor action. These anticipatory responses tend to be complex, involving many synergistic muscle groups. Anticipatory responses must be learned, but eventually they operate automatically, being triggered by specific intended movements. The postural system is also equipped with stereotyped response patterns that are rapidly corrected for unexpected perturbations. Some of these responses are innate, while others have to be acquired by motor learning that involves the cerebellum. These responses are characteristically driven by immediate feedback from visual, vestibular, and somatosensory information. In the past, posture might have been explained by the parallel action of involuntary reflexes controlled at relatively low levels of the nervous system. Today we recognize that postural control is complex and context-dependent and that all levels of the nervous system must be examined to account for this complexity.
- Systematic investigation of posture began with experiments on four-legged animals, notably the decerebrate cat, in which cerebral control of the brain stem is disconnected by a transection at a midbrain level (see Box 37-1). This preparation was devised by Charles Sherrington at the beginning of the twentieth century to study spinal reflexes isolated from higher levels of motor control. In a series of classic experiments on decerebrate cats and dogs early in the twentieth century, Sherrington, Rudolf Magnus, and other physiologists uncovered a variety of stretch reflexes because these reflexes are exaggerated in the decerebrate animal (Chapter 36). When the cut is between the superior and inferior colliculi, contraction of extensor and other antigravity muscles is intensified, producing what is called decerebrate rigidity. The rigidity may be great enough to allow the animal to stand unsupported. When the sensory nerves from the limbs of decerebrate animals are cut, the enhanced tone of antigravity muscles collapses dramatically, revealing that afferent signals from muscle are crucial to efferent motor control. Sherrington went so far as to suggest that rigidity of extensor muscles in the face of gravity represents the foundation of postural control expressed in “grotesque form,” although he knew that there was more to postural control than this. When Magnus and his colleagues transected the brain at higher levels they observed a hierarchy of automated responses to postural disturbance—ranging from head and neck reflexes to the so-called righting reflexes—that are not possible in the midbrain decerebrate preparation. Magnus concluded that postural control could be explained by the simple summation of these reflexes. Postural reflexes do not fully account for postural control during skilled, purposeful movements, however. For example, the simple act of reaching out to grasp an object can destabilize balance unless precisely timed compensatory action is initiated before the arm is extended. Thus a system is needed to generate anticipatory responses. In addition, a wide range of automatic response patterns must be available to cope with any unexpected disturbances. Finally, because postural control must be integrated with voluntary movement, postural control systems must be capable of adaptive learning. Thus the current view is that postural control involves the behaviorally meaningful integration of many different neural systems, including those associated with cognition.
- Figure 17.1. Local circuit neurons that supply the medial region of the ventral horn are situated medially in the intermediate zone of the spinal cord gray matter and have axons that extend over a number of spinal cord segments and terminate bilaterally. In contrast, local circuit neurons that supply the lateral parts of the ventral horn are located more laterally, have axons that extend over a few spinal cord segments, and terminate only on the same side of the cord. Descending pathways that contact the medial parts of the spinal cord gray matter are involved primarily in the control of posture; those that contact the lateral parts are involved in the fine control of the distal extremities Figure 17.2. Descending projections from the brainstem to the spinal cord. Pathways that influence motor neurons in the medial part of the ventral horn originate in the reticular formation, vestibular nucleus, and superior colliculus. Those that influence motor neurons that control the arm muscles originate in the red nucleus and terminate in more lateral parts of the ventral horn. Upper Motor Neuron Control of the Brainstem and Spinal Cord Overview The axons of upper motor neurons descend from higher centers to influence the local circuits in the brainstem and spinal cord that organize movements. The sources of these upper motor neuron pathways include several brainstem centers and a number of cortical areas in the frontal lobe. The motor control centers in the brainstem are especially important in ongoing postural control. Each center has a distinct influence. Two of these centers, the vestibular nuclear complex and the reticular formation, have widespread effects on body position. Another brainstem center, the red nucleus, controls movements of the arms; also in the brainstem, the superior colliculus contains upper motor neurons that initiate orienting movements of the head and eyes. The motor and “premotor” areas of the frontal lobe, in contrast, are responsible for the planning and precise control of complex sequences of voluntary movements. Most upper motor neurons, regardless of their source, influence the generation of movements by directly affecting the activity of the local organizing circuits in the brainstem and spinal cord (see Chapter 16 ). Upper motor neurons in the cortex also control movement indirectly, via pathways that project to the brainstem motor control centers, which, in turn, project to the local organizing circuits in the brainstem and cord. A major function of these indirect pathways is to maintain the body's posture during cortically initiated voluntary movements Descending Control of Spinal Cord Circuitry: General Information Some insight into the functions of the different sources of the upper motor neurons is provided by the way the lower motor neurons and local circuit neurons—the ultimate targets of the upper motor neurons—are arranged within the spinal cord. As described in Chapter 16 , lower motor neurons in the ventral horn of the spinal cord are organized in a somatotopic fashion: The most medial part of the ventral horn contains lower motor neuron pools that innervate axial muscles or proximal muscles of the limbs, whereas the more lateral parts contain lower motor neurons that innervate the distal muscles of the limbs. The local circuit neurons, which lie primarily in the intermediate zone of the spinal cord and supply much of the direct input to the lower motor neurons, are also topographically arranged. Thus, the medial region of the intermediate zone of the spinal cord gray matter contains the local circuit neurons that synapse withlower motor neurons in the medial part of the ventral horn, whereas the lateral regions of the intermediate zone contain local neurons that synapse primarily with lower motor neurons in the lateral ventral horn. The patterns of connections made by local circuit neurons in the medial region of the intermediate zone are different from the patterns made by those in the lateral region, and these differences are related to their respective functions ( Figure 17.1 ). The medial local circuit neurons, which supply the lower motorneurons in the medial ventral horn, have axons that project to many spinal cord segments; indeed, some project to targets along the entire length of the cord. Moreover, many of these local circuit neurons also have axonal branches that cross the midline in the commissure of the spinal cord to innervate lower motorneurons in the medial part of the contralateral hemicord. This arrangement ensures that groups of axial muscles on both sides of the body act in concert to maintain and adjust posture. In contrast, local circuit neurons in the lateral region of the intermediate zone have shorter axons that typically extend fewer than five segments and are predominantly ipsilateral. This more restricted pattern of connectivity underlies the finer and more differentiated control that is exerted over the muscles of the distal extremities, such as that required for the independent movement of individual fingers during manipulative tasks. Differences in the way upper motor neuron pathways from the cortex and brainstem terminate in the spinal cord conform to these functional distinctions between the local circuits that organize the activity of axial and distal muscle groups. Thus, most upper motor neurons that project to the medial part of the ventral horn also project to the medial region of the intermediate zone; the axons of the neurons have collateral branches that terminate over many spinal cord segments, reaching medial cell groups on both sides of the spinal cord. The sources of these projections are primarily the vestibular nuclei and the reticular formation (see next section); as their terminal zones in the medial spinal cord gray matter suggest, they are concerned primarily with postural mechanisms ( Figure 17.2 ). In contrast, descending axons from the motor cortex generally terminate in lateral parts of the spinal cord gray matter and have terminal fields that are restricted to only a few spinal cord segments. These corticospinal pathways are primarily concerned with precise movements involving more distal parts of the limbs. Two additional brainstem structures, the superior colliculus and the red nucleus, also contribute upper motor neuron pathways to the spinal cord. The axons arising from the superior colliculus project to medial cell groups in the cervical cord, where they influence the lower motor neuron circuits that control axial musculature of the neck (see Figure 17.2 ). These projections are particularly important in generating orienting movements of the head (the role of the superior colliculus in the generation of head and eye movements is covered in detail in Chapter 20 ). The red nucleus projections are also limited to the cervical level of the cord, but these terminate in lateral regions of the ventral horn and intermediate zone (see Figure 17.2 ). In humans, the axons arising from the red nucleuscontrol primarily the proximal muscles of the arm. The limited distribution of rubrospinal projections may seem surprising, given the large size of the red nucleus in humans ( rubro - means red; the adjective is derived from the rich capillary bed that gives the nucleus a reddish color in fresh tissue). In fact, the bulk of the red nucleus in humans is a subdivision that does not project to the spinal cord at all, but relays information from the cortex to the cerebellum (see Chapter 19 ).
- Figure 17.3. The location of the reticular formation in relation to some other major landmarks at different levels of the brainstem. Neurons in the reticular formation are scattered among the axon bundles that course through the medial portion of the midbrain, pons, and medulla. Motor Control Centers in the Brainstem: Upper Motor Neurons That Maintain Balance and Posture As described in Chapter 14 , the vestibular nuclei are the major destination of the axons that form the vestibular division of the eighth cranial nerve; as such, they receive sensory information from the semicircular canals and the otolith organs that specifies the position and angular acceleration of the head. Many of the cells in the vestibular nuclei that receive this information are upper motor neurons with descending axons that terminate in the medial region of the spinal cord gray matter, although some extend more laterally to contact the neurons that control the proximal muscles of the limbs. The projections from the vestibular nuclei that control axial muscles and those that influence proximal limb muscles originate from different cells and take different routes (called the medial and lateral vestibulospinal tracts). The reticular formation is a complicated network of circuits located in the core of the brainstem and extending from the rostral midbrain to the caudal medulla ( Figure 17.3 ). Unlike the well-defined sensory and motor nuclei of the cranial nerves, the reticular formation comprises clusters of neurons scattered among a welter of interdigitating axon bundles; it is therefore difficult to subdivide anatomically. The neurons within the reticular formation have a variety of functions, including cardiovascular and respiratory control (see Chapter 21 ), governance of myriad sensory motor reflexes (see Chapter 16 ), the organization of eye movements (see Chapter 20 ), regulation of sleep and wakefulness (see Chapter 28 ), and, most important for present purposes, motor control. The descending motor control pathways from the reticular formation to the spinal cord are similar to those of the vestibular nuclei: They terminate primarily in the medial parts of the gray matter where they influence the local circuit neurons that coordinate axial and proximal limb muscles (see Figure 17.2 ). Both the vestibular nuclei and the reticular formation provide information to the spinal cord that maintains posture in response to environmental (or self-induced) disturbances of body position and stability. As expected, the vestibular nuclei make adjustments in posture and equilibrium in response to information from the inner ear. Direct projections from the vestibular nuclei to the spinal cord ensure a rapid compensatory response to any postural instability detected by the inner ear (see Chapter 14 ). In contrast, the motor centers in the reticular formation are controlled largely by other motor centers in the cortex or brainstem. The relevant neurons in the reticular formation initiate adjustments that stabilize posture during ongoing movements.
- Figure 17.6. Direct and indirect pathways from the motor cortex to the lateral (A) and medial (B) gray matter of the spinal cord. Neurons in the motor cortex that supply the lateral part of the ventral horn also terminate on neurons in the red nucleus. Neurons in the motor cortex that supply the medial part of the ventral horn also terminate on neurons in the reticular formation. Thus, the motor cortex has both direct and indirect routes by which it can influence the activity of spinal cord neurons. The results of this experiment can be understood in terms of the fact that the upper motor neurons in the motor cortex influence the spinal cord circuits by two routes: direct projections to the spinal cord and indirect projections to brainstem centers under consideration here that in turn project to the spinal cord (Figure 17.6). The reticular formation is one of the major destinations of these latter projections from the motor cortex; thus, cortical upper motorneurons initiate both the reaching movement of the forepaw and also the postural adjustments in the other limbs necessary to maintain body stability. The forepaw movement is initiated by the direct pathway from the cortex to the spinal cord (and possibly by the red nucleus as well), whereas the postural adjustments are mediated via pathways from the motor cortex that reach the spinal cord indirectly, after an intervening relay in the reticular formation (the corticoreticulospinal pathway). Further evidence for the contrasting functions of the direct and indirect pathways from the motor cortex and brainstem to the spinal cord has come from experiments carried out by the Dutch neurobiologist Hans Kuypers, who examined the behavior of rhesus monkeys that had the direct pathway transected at the level of the medulla, leaving the indirect descending upper motor neuron pathways to the spinal cord via the brainstem centers intact. Immediately after the surgery, the animals were able to use axial and proximal muscles to stand, walk, run, and climb, but they had great difficulty using the distal parts of their limbs (especially their hands) independently of other body movements. For example, the monkeys could cling to the cage but were unable to reach toward and pick up food with their fingers; rather, they used the entire arm to sweep the food toward them. After several weeks, the animals recovered some independent use of their hands and were again able to pick up objects of interest, but this action still involved the concerted closure of all of the fingers. The ability to make independent, fractionated movements of the fingers, as in opposing the movements of the fingers and thumb to pick up an object, never returned. These observations show that following damage to the direct corticospinal pathway at the level of the medulla, the indirect projections from themotor cortex via the brainstem centers (or from brainstem centers alone) are capable of sustaining motor behavior that involves primarily the use of proximal muscles. In contrast, the direct projections from the motor cortex to the spinal cord enable the speed and agility of movements, providing a higher degree of precision in fractionated finger movements than is possible using the indirect pathways alone. Selective damage to the corticospinal tract (i.e., the direct pathway) in humans is rarely seen in the clinic. Nonetheless, this evidence in nonhuman primates showing that direct projections from the cortex to the spinal cord are essential for the performance of discrete finger movements helps explain the limited recovery in humans after damage to the motor cortex or to the internal capsule. Immediately after such an injury, such patients are typically paralyzed. With time, however, some ability to perform voluntary movements reappears. These movements, which are presumably mediated by the brainstem centers, are crude for the most part, and the ability to perform discrete finger movements such as those required for writing, typing, or buttoning typically remains impaired